

Анотація
Зростає усвідомлення викликів, пов’язаних із забезпеченням гарним харчуванням усіх живих істот без винятку без необхідності досягнення компромісів з довкіллям. Це особливо вірно для аквакультури, враховуючи залежність аквакультури від вилучення морських живих ресурсів як ключових інгредієнтів для комбікормів, одночасно виступаючи постачальником таких ключових поживних речовин як омега-3 довголанцюгові поліненасичені жирні кислоти. У цьому оглядові обговорюється прогрес на шляху переходу від рибної олії океанічного походження як інгредієнту кормів з наголосом на нових трансгенних рослинних джерелах цих жирних кислот. Особливу вагу приділено фазі “перевірки” цього процесу, під час якої олія з ГМ-рослин використовувалась як замінник справжньої риб’ячої олії у кормах для гідробіонтів. Належним чином береться до уваги демонстрація “реального” потенціялу шляхом «польових» випробувань ГМ. У сукупності статус цих нових рослинних джерел риб’ячого жиру омега-3 підтверджує появу нової хвилі рослинних біотехнологічних продуктів через 25 років після введення в оборот стійких до гербіцидів рослин, і демонструє силу ГМ сільського господарства для сприяння продовольчій безпеці та роботі в межах планети.
Вступ
Користь для здоров’я, пов’язана з дієтою, що містить жирну рибу, багату поліненасиченими омега-3 довголанцюговими жирними кислотами (omega-3 LC- PUFAs), тепер добре доведена, і вона пов’язана з регулярним споживанням цих жирних кислот та зниженням ризику серцево-судинних захворювань та інфаркта міокарда[1]. Однак багато факторів поєднались, щоб зробити цю, очевидно, просту рекомендацію менш легкою для виконання, і серед таких чинників – невпинне зростання населення планети, яке висуває зростаючі вимоги до обмежених ресурсів нашої планети, забруднення морського середовища та поступова зміна клімату [2,3]. Можливо, парадоксально, але зростання аквакультури як домінуючої системи вирощування риби для споживання людиною змінило динаміку пропозиції та попиту на омега-3, створюючи додаткове навантаження на обмежені світові запаси риби. [2,3]. Як обговорюється нижче, спроби зробити аквакультуру більш сталою призвели до зниження харчової цінності кінцевого продукту [4]. У цьому сенсі включення олій наземних рослин, основної альтернативи риб’ячому жиру в Європі, в аквакорми призвело до помітного зниження рівня омега-3 LC-PUFAs у спожитих харчах, що супроводжується підвищенням рівня коротколанцюгових жирних кислот, характерних для наземних систем. За останні кілька десятиліть давно усталені схеми харчування радикально змінилися, аж до того, що метаболічні захворювання, спричинені надмірним споживанням продуктів, багатих калоріями, але з низьким вмістом поживних речовин, вражають дедалі більшу кількість людей, які страждають від недоїдання через недостатнє харчування. Одночасно з цим зростаюче усвідомлення того, що глобальні моделі споживання їжі виходять за рамки можливостей планети, призвело до радикальних змін у тому, як ми харчуємось, особливо через значне зменшення споживання білка (наземного) тваринного походження та супутнє збільшення в харчуванні рослинних білків [5]. Цікаво, що також рекомендується збільшене споживання риби (через поживні переваги омега-3 тощо), але така порада пов’язана з невід’ємною напругою, не в останню чергу щодо сталості світових запасів риби [2,5,6,7].
Біотехнологія рослин завжди прагнула принести користь суспільству, навіть якщо іноді це не було очевидно [8]. Перша хвиля ГМ-культур, розроблена в 1990-х роках, була зосереджена на забезпеченні так званих вхідних властивостей, які допомагають фермерам (через толерантність до гербіцидів та/або стійкість до травоїдних тварин), але не приносять прямої користі для споживача чи підвищення поживності (хоча вони забезпечують меншу залежність від хімічних речовин). Друга хвиля ГМ-культур, вже з урахуванням вихідних властивостей в частині покращеного вмісту поживних речовин, лише зараз починає з’являтися, тобто через 25 років після появи сортів з орієнтацією на вхідні ознаки; численні та різноманітні причини цього нещодавно були розглянуті нами та іншими авторами [8,9]. На передньому краї цих нових ГМ-культур є рослини, які були сконструйовані для накопичення омега-3 LC-PUFAs у їхніх насіннєвих оліях, забезпечуючи нове наземне джерело цих важливих жирних кислот, незалежне від морських запасів [10] – такі рослинні джерела “oмега-3 риб’ячої олії” можуть використовуватись або безпосереднього вживання людиною або як інгредієнти корму для тварин (як гідробіонтів, так і суходільних), приносячи користь такими різними шляхами [11].
Омега-3 LC-PUFAs, ойкозопентаєнова кислота (EPA; 20:5Δ5,8,11,14,17) та докозопентаєнова кислота (DHA; 22:6Δ4,7,10,13,16,19) містяться у риб’ячій олії та майже виключно обмежені присутністю лише у водних харчових ланцюгах [2,3]. Основними живими істотами, що продукують EPA та DHA у морських екосистемах, є мікроводорості та інший фітопланктон, з жирними кислотами, що накопичуються на багатьох трофічних рівнях вище цих організмів [2]. Морські та лососеві види риб мають невелику або зовсім не мають ендогенної здатності синтезувати ці жирні кислоти, натомість покладаючись на споживання їх з їжею. Жоден вид покритонасінних рослин не має здатності синтезувати EPA та DHA, хоча С18 – попередники поліненасичених жирних кислот є всюдисущими в Царстві Рослин [4]. Починаючи з 90—х років здійснювались значні зусилля у напрямі конструювання рослин для акумуляції EPA та/або DHA як альтернативи вилученню їх з морських організмів [8–10]. Продукування омега-3 риб’ячого жиру в трансгенних рослинах представляє собою один із небагатьох проєктів, який спочатку перейшов від фази відкриття до застосування, і є значним досягненням для рослинної біотехнології. Цей короткий огляд буде зосереджено на нещодавніх прикладах масштабування та перекладу, включно з демонстрацією ефективності під час випробувань аквакормів. Аспекти молекулярної біології та генетичних інструментів, які використовувалися для отримання цих трансгенних рослин, нещодавно розглядалися в інших місцях і тому тут обговорюються лише коротко.
Трансгенні рослини як платформа для синтезу EPA + DHA
Незважаючи на оманливо просту мету, ефективний синтез неприродних для суходільних рослин жирних кислот EPA та DHA виявився складним для досягнення, особливо коли акумуляція обмежувалася зберіганням триацилгліцеринів через специфічні для насіння промотори [12–14]. Частково це було пов’язано з відносною складністю біосинтетичного шляху, який вимагав узгодженої експресії мінімум п’яти гетерологічних генів, що представляло собою одну з найскладніших метаболічних конструкцій, застосованих у трансгенних рослинах [10]. Однак завдяки застосуванню ітераційного підходу до визначення оптимальної комбінації генів і регуляторних елементів було досягнуто значного прогресу у двох видів- господарів рижія та [12,15,16] ріпака [17,18]. Цікаво, що також стало очевидним, що ендогенний метаболізм відіграє ключову роль у забезпеченні ефективного відновлення шляху біосинтезу омега-3 LC-PUFA, і ця притаманна здатність різниться між видами. Наприклад, успішний синтез EPA + DHA в Arabidopsis (гусимці) або Camelina (рижії) через так званий альтернативний шлях [19,20] не був повторений в арахісі [21]. Навіть у випадку ефективних сортів олійних культур, таких як ріпак та рижій, існують значні відмінності між видами, наприклад, дуже подібний набір генів і регуляторних елементів використовувався для керування синтезом EPA + DHA, в результаті чого у Camelina було ~20% EPA + DHA, але лише 8% EPA і 1% DHA в ріпакові. Це може свідчити про додаткові рівні контролю, які можуть включати місце інтеграції трансгену, посттранскрипційну регуляцію, обмін ферментів і каналізацію субстрату. Незалежно від цих відкритих питань, очевидно, що олійні насіння Brassicacea (капустяні) є чудовими господарями для трансгенного накопичення омега-3 LC-PUFA (узагальнено в Таблиці 1), а олії, отримані з цих культур, використовувалися в багатьох випробуваннях кормів для гідробіонтів. Важливо, що ефективність цих культур у реальних умовах була продемонстрована польовими випробуваннями в багатьох місцях на різних континентах [22-24]. Нещодавно редагування генів було використано в поєднанні з трансгенезом для посилення накопичення EPA + DHA у рижії через інактивацію конкуруючого ендогенного шляху [25].
| Таблиця 1 | |||||
| Резюме ГМ капустяних культур, успішно створених для накопичення EPA та/або DHA. Рівні окремих жирних кислот наведено разом із загальною кількістю омега-3 LC-PUFAs. У більшості всіх успішних технологічних рішень використовувався шлях аеробної десатурази та елонгази, хоча один приклад наведено з використанням анаеробної полікетидсинтазоподібної системи (PKS-подібна система), отриманої з мікробних джерел. Юудь ласка, дивіться [2] and [10] для більшої деталізації щодо шляхів. | |||||
| Культура | Шлях | Профіль Omega-3 | Загальний вміст EPA + DHA | Стан комерціалізації | Посилання |
| Рижій Рижій
Ріпак Ріпак Ріпак |
аеробний – Rothamsted аеробний – CSIRO
аеробний – BASF аеробний – Nuseed/CSIRO анаеробний (PKS-подібна) – Dow Agrosciences |
10% EPA, 10% DHA
2% EPA, 12% DHA 7% EPA, 1% DHA
1% EPA, 10% DHA
4% DHA |
20%
14% <10%
12%
<5% |
Дорегуляторна фаза Дорегуляторна фаза Схвалено для комер-ційного вирощування
Схвалено для комер-ційного вирощування Невідомий |
[12]
[15] [18]
[17●] [38] |
Новітні олії в кормах для гіробіонтів – перевірка в експериментах з годівлею риби
Одним з ключових аспектів досягнення успішної трансляції біотехнологічних досліджень, описаних вище, у масштабоване виробництво, є перевірка цих нових олій як ефективних замінників риб’ячого жиру в раціонах, які використовуються в аквакультурі (представлено на рис. 1).
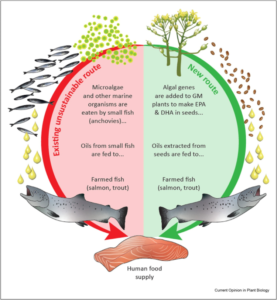
Схематичне зображення потреби в омега-3 риб’ячому жирі в морській аквакультурі та того, як ГМ-жирні кислоти рослинного походження можна використовувати як заміну для оптимального харчування риби та людини.
На сьогоднішній день більшість досліджень риби з використанням жирів, похідних з ГМ-джерел, багатих на n-3 LC-PUFA, були переважно зосереджені на сьомзі (Salmo salar). Це значною мірою пов’язано з тим фактом, що лососеві є основним споживачем екстрагованого риб’ячого жиру, враховуючи енергоємні корми, необхідні для вирощування, а також високу цінність кінцевої продукції [4,26]. Тому вкрай важливо досліджувати потенційні наслідки використання таких замінників. Отже, олії, отримані з багатих на омега-3 Camelina [12,23] та Canola (Event NS-B50027-4) [17,18] були успішно випробувані на декількох стадіях життєвого циклу гідробіонтів. Прісноводна молодь, яку годували 4 або 8% ГМ ріпакової олії з високим вмістом n-3 кислот (9,3% омега LC-PUFA), продемонстрували ріст і виживання, подібні до тих особин, яких годували дієтою, що містила риб’ячий жир як основне харчове джерело ліпідів, за культивування при 12 або 160C [27]. У морській воді не було виявлено різниці в рості смолтів, яких годували олією рижія, багатою на EPA або EPA + DHA (23,9% і 16,7% омега-3 LC-PUFA, відповідно), порівняно з тими, яких годували дієтою, багатою риб’ячим жиром [28–31]. Крім того, усі вищезазначені випробування показали, що риба, яку годували жирами ГМ- походження, мала підвищений рівень n-3 LC-PUFA у м’ясі порівняно з комерційним контрольним раціоном або кормом, що містив рижія дикого типу, а також знижене співвідношення омега-жирних кислот (омега -6: омега-3). Наприклад, лосось, якого годували дієтою з високим вмістом омега-3 LC-PUFA з олії рижія, накопичував майже вдвічі більше цих корисних для здоров’я жирних кислот, ніж лосось, якого годували риб’ячим жиром [31]. Очевидно, що ці підвищені рівні можуть сприяти подоланню розриву між попитом і пропозицією та допомогти задовольнити потребу в споживанні 500 мг/день для омега-3 LC-PUFA для досягнення оптимального здоров’я серця [1,2,4]. Підвищення рівня EPA + DHA в м’ясі було загальним висновком інших досліджень, у яких кісткових риб годували омега-3 LC- PUFA з рослинних олій, таких як такі жирні види, як райдужний пструг (Oncorhynchus mykiss) [32], а також дорада (Sparus aurata) [33] та лаврак (Dicentrarchus labrax) [34]. На додаток до переваг, які використання цих олій може мати для споживачів, використання рослинних олій, багатих на омега-3 LC-PUFA, також може мати додаткові переваги для риб, такі як зменшення інфільтрації жиру в печінці та площі гепатоцитів порівняно з рибою, яку годують кормами, що містять рижія дикого типу [35]. Мабуть, найважливіше відзначити те, що жодне з проведених на сьогоднішній день випробувань не повідомило про будь-який несприятливий вплив на рибу, яку годували цими новими оліями, що демонструє потенціал для використання цієї ключової сировини [27-30,31-36]. Поки що дослідження були зосереджені на використанні лише олії, тоді як макуха ще не була перевірена як альтернативне джерело білка. Для концентрації білкової фракції може знадобитися додаткова обробка насіння, наприклад лущення або екстракція розчинником, що, у свою чергу, збільшить ціну, а також негативний вплив процесу виробництва продукту на навколишнє середовище.
Наступні кроки та нові підходи
Перевірка цих нових олій як безпечних і ефективних замінників риб’ячого жиру океанічного походження стала ключовим кроком у зниженні ризиків цієї технології та сприянні її виходу на ринок. Крім того, підвищена обізнаність щодо питань, пов’язаних із сталістю та впливом на навколишнє середовище [5,11,26] додала імпульсу до комерціалізації. Чудово, що два сорти каноли, розроблені BASF/Cargill і Nuseed/CSIRO, були дерегульовані (виведені з процедури регулювання) USDA, і тепер їх можна вирощувати у великих масштабах [18]. Ці події являють собою найскладнішу метаболічну інженерію в рослинах, схвалену для комерційного вирощування на сьогоднішній день, хоча ці розробки є кульмінацією 25 років фундаментальних досліджень з подальшою узгодженою зосередженістю на розробці [8]. Одночасно олія з рижія широко використовувалась у дослідженнях на людях, що підтверджує корисність цієї олії як у непрямому (тобто через тварин), так і в прямому харчуванні людини [37]. Існують також спроби поєднати низку різних ознак для отримання багатофункціональної культури, адаптованої до потреб, напр. аквакультури [11], що поєднує властивість EPA+DHA з іншими високоцінними компонентами раціонів для аквакорму, такими як антиоксидант і пігмент астаксантин, який можна виробляти в ряді різних трансгенних господарів і щодо якого було незалежно підтверджено як нового інгредієнту у дослідах з годування риб [39,40]. Подібним чином можна покращити амінокислотний склад і запасні білки насіння, щоб зробити отриманий білковий склад більш придатним для харчування риб (наприклад, шляхом включення похідного цистеїну, таурину), як це було зроблено в трансгенній сої [40]. Не менш важливо, що можна використовувати редагування генів для видалення таких сполук, як глюкозинолати, які можуть діяти як стримуючий фактор для годування деяких видів риб [41,42]. На цих засадах можна передбачити індивідуальний ідеотип культури, спеціально створеної для задоволення харчових потреб як риби, так і людини як прямого споживача, а також такого, що відповідатиме вимогам оперування в межах планети та залишатиме менший екологічний слід [11,43].
Висновки
Нещодавній аналіз підтверджує значні варіяції між попитом omega-3 довголанцюговіin PUFAs та доступністю цих речовин у наших океанах, що зараз оцінюється у ~30% від рекомендованого рівня EPA та DHA для населення планети [44]. Це підкреслює нагальну потребу в альтернативних поставках цих важливих жирних кислот, не в останню чергу враховуючи їх роль у захисті здоров’я від серцево-судинних захворювань та інших метаболічних патологій, які зараз псують життя багатьох мільйонів людей. На додаток до сталого синтезу EPA & DHA з використанням трансгенних рослин, зараз створено низку мікробіальних (переважно водорості) систем [огляд наведено у [45]], і вони також підлягають валідації цих новітніх олій таким самим шляхом в експериментах годування риби [46]. Однією з переваг використання омега-3 PUFA, отриманих із водоростей, є те, що ці штами зазвичай не є ГМО, що робить їх схвалення регуляторами та прийняття споживачами відносно простими. Проте широкомасштабне вирощування штамів водоростей залишається проблемою з точки зору економіки та інфраструктури. Більше того, майже всі комерційні водоростеві платформи покладаються на гетеротрофний ріст (тобто культивуються з використанням екзогенного джерела вуглецю, який сам незмінно походить із рослин, наприклад, цукрової тростини). Це не завжди повністю відображається у заявах щодо сталості, і буде цікаво провести аналіз життєвого циклу різних виробничих систем, щоб дозволити споживачеві зробити усвідомлений вибір щодо екологічного сліду. Інші джерела LC-PUFA, такі як масло криля, зараз є основними інгредієнтами кормів для аквакорму, хоча їхнє включення зазвичай обмежене в основному через високу ціну [2]. Іншим підходом до підвищення рівня цих корисних для здоров’я жирних кислот у рибі без впливу на рибальство є використання фінішних дієт з високим вмістом LC-PUFA [47]. Хоча новітня платформа з отримання олій, що описана вище, є ГМ з відповідним регуляторним обтяженням [8,9], вони мають переваги у використанні вже існуючох інфаструктури та ноу-хау, що лежать в основі сучасного сільського господарства, що дозволяє швидке масштабування та інкорпорацію у виробничі цикли. Розширення використання рослинних кормів і харчових продуктів буде мати важливе значення для роботи в межах планети, і разом ми повинні прийняти інший підхід до забезпечення оптимального харчування для всіх – рослинна біотехнологія може відіграти ключову роль у цьому. Таким чином, важливо враховувати всі етичні аспекти використання ГМ інгредієнтів, включно з впливом того, що ці інновації не буде сприйнято/ухвалено [48].
Посилання
-
Marine omega-3 (N-3) fatty acids for cardiovascular health: an update for 2020Int J Mol Sci, 21 (2020), p. 1362, 10.3390/ijms21041362
- 2•
Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: bridging the gap between supply and demandNutrients, 11 (2019), p. 89, 10.3390/nu11010089
-
A comprehensive review comparing different new sources of EPA and DHA, including algal and transgenic plants
- 3
Recent advances in omega-3: health benefits, sources, products and bioavailabilityNutrients, 6 (2014), pp. 3727-3733, 10.3390/nu6093727
- 4
Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015Sci Rep, 6 (2016), Article 21892
- 5••
Food in the Anthropocene: the EAT–Lancet Commission on healthy diets from sustainable food systemsLancet, 393 (2019), pp. 447-492
-
A ground-breaking analysis of the challenges of feeding the global population within so-called planetary boundaries. The study recommends a shift to a diet which is predominantly plant-based.
- 6
Avoiding the ecological limits of forage fish for fed aquacultureNat Sustain, 1 (2018), pp. 298-303, 10.1038/s41893-018-0077-1
- 7
Global adoption of novel aquaculture feeds could substantially reduce forage fish demand by 2030Nat Food, 1 (2020), pp. 301-308, 10.1038/s43016-020-0078-x
- 8
The challenges of delivering genetically modified crops with nutritional enhancement traitsNative Plants, 5 (2019), pp. 563-567, 10.1038/s41477-019-0430-z
- 9
Multiplying the efficiency and impact of biofortification through metabolic engineeringNat Commun, 11 (2020), p. 5203, 10.1038/s41467-020-19020-4
- 10
Transgenic plants as a sustainable, terrestrial source of fish oilsEur J Lipid Sci Technol, 117 (2015), pp. 1317-1324, 10.1002/ejlt.201400452
- 11•
Agriculture can help aquaculture become greenerNat Food, 1 (2020), pp. 680-683, 10.1038/s43016-020-00182-9
-
This review defines the issues facing aquaculture, balancing the laudable goal of becoming more sustainable against the challenge of maintaining the nutritional benefits from consuming fish. A wider role for agriculture and plant biotechnology is proposed.
- 12
Successful high-level accumulation of fish oil omega-3 long-chain polyunsaturated fatty acids in a transgenic oilseed cropPlant J, 77 (2014), pp. 198-208, 10.1111/tpj.12378
- 13
Metabolic engineering plant seeds with fish oil-like levels of DHAPLoS One, 7 (2012), Article e49165, 10.1371/journal.pone.0049165
- 14
Synthetic redesign of plant lipid metabolismPlant J, 87 (2016), pp. 76-86, 10.1111/tpj.13172
- 15
Metabolic engineering Camelina sativa with fish oil-like levels of DHAPLoS One, 9 (2014), Article e85061, 10.1371/journal.pone.0085061
- 16
Characterization of oilseed lipids from “DHA-producing Camelina sativa”: a new transformed land plant containing long-chain omega-3 oilsNutrients, 6 (2014), pp. 776-789, 10.3390/nu6020776
- 17•
Development of a Brassica napus (canola) crop containing fish oil-like levels of DHA in the seed oilFront Plant Sci, 11 (2020), p. 727, 10.3389/fpls.2020.00727
-
This paper describes the molecular and agronomic performance of the specific Canola event taken through the regulatory approval process.
- 18
Update on GM canola crops as novel sources of omega-3 fish oilsPlant Biotechnol J, 17 (2019), pp. 703-705, 10.1111/pbi.13045
- 19
Production of very long chain polyunsaturated omega-3 and omega-6 fatty acids in plantsNat Biotechnol, 22 (2004), pp. 739-745, 10.1038/nbt972
- 20
An alternative pathway for the effective production of the omega-3 long-chain polyunsaturates EPA and ETA in transgenic oilseedsPlant Biotechnology Journal, 13 (2015), pp. 1264-1275, 10.1111/pbi.123282015
- 21
Production of eicosapentaenoic acid (EPA, 20:5n-3) in transgenic peanut (Arachis hypogaea L.) through the alternative Δ8-desaturase pathwayMol Biol Rep, 46 (2019), pp. 333-342, 10.1007/s11033-018-4476-1
- 22
Tailoring seed oil composition in the real world: optimising omega-3 long chain polyunsaturated fatty acid accumulation in transgenic Camelina sativaSci Rep, 7 (2017), p. 6570, 10.1038/s41598-017-06838-0
- 23
High level accumulation of EPA and DHA in field-grown transgenic Camelina – a multi-territory evaluation of TAG accumulation and heterogeneityPlant Biotechnology Journal, 18 (2020), pp. 2280-2291, 10.1111/pbi.13385
- 24•
Using field evaluation and systematic iteration to rationalize the accumulation of omega-3 long-chain polyunsaturated fatty acids in transgenic Camelina sativaPlant Biotechnology Journal (2022), 10.1111/pbi.13867
-
A significant study incorporating field release to determine the performance of new iterations of transgenic Camelina making EPA and DHA. The need to include real-world performance in the “design-build-test-learn” cycle is emphasised.
- 25••
Enhancing the accumulation of eicosapentaenoic acid and docosahexaenoic acid in transgenic Camelina through the CRISPR-Cas9 inactivation of the competing FAE1 pathwayPlant Biotechnology Journal (2022), 10.1111/pbi.13876
-
The first example of a GM and GE stack undergoing field release. Specifically, gene editing was used to remove a competing pathway, enhancing the transgenic accumulation of EPA and DHA in Camelina.
- 26
A 20-year retrospective review of global aquacultureNature, 591 (2021), pp. 551-563, 10.1038/s41586-021-03308-6
- 27•
n-3 Canola oil effectively replaces fish oil as a new safe dietary source of DHA in feed for juvenile Atlantic salmonBr J Nutr, 122 (2019), pp. 1329-1345, 10.1017/S0007114519002356
-
The first study on the performance of the Nuseed DHA Canola oil in aquafeed studies on salmon.
- 28
A nutritionally-enhanced oil from transgenic Camelina sativa effectively replaces fish oil as a source of eicosapentaenoic acid for fishSci Rep, 5 (2015), p. 8104, 10.1038/srep08104
- 29
Evaluation of a high-EPA oil from transgenic Camelina sativa in feeds for Atlantic salmon (Salmo salar L.): effects on tissue fatty acid composition, histology and gene expressionAquaculture, 444 (2015), pp. 1-12, 10.1016/j.aquaculture.2015.03.020
- 30
Nutritional evaluation of an EPA-DHA oil from transgenic camelina sativa in feeds for post-smolt atlantic salmon (Salmo salar L.)PLoS One, 11 (2016), Article e0159934, 10.1371/journal.pone.0159934
- 31
An oil containing EPA and DHA from transgenic Camelina sativa to replace marine fish oil in feeds for Atlantic salmon (Salmo salar L.): effects on intestinal transcriptome, histology, tissue fatty acid profiles and plasma biochemistryPLoS One, 12 (2017), Article e0175415, 10.1371/journal.pone.0175415
- 32
Transgenic camelina oil is an effective source of eicosapentaenoic acid and docosahexaenoic acid in diets for farmed rainbow trout, in terms of growth, tissue fatty acid content, and fillet sensory propertiesJ World Aquacult Soc, 52 (2021), pp. 961-986
- 33
Replacement of Marine Fish Oil with de novo Omega-3 Oils from Transgenic Camelina sativa in Feeds for Gilthead Sea Bream (Sparus aurata L.)Lipids, 51 (2016), pp. 1171-1191, 10.1007/s11745-016-4191-4
- 34
Oil from transgenic Camelina sativa as a source of EPA and DHA in feed for European sea bass (Dicentrarchus labrax L.)Aquaculture, 530 (2021), Article 735759, 10.1016/j.aquaculture.2020.735759
- 35
Oil from transgenic Camelina sativa containing over 25 % n-3 long-chain PUFA as the major lipid source in feed for Atlantic salmon (Salmo salar)Br J Nutr, 119 (2018), pp. 1378-1392, 10.1017/S0007114518001125
- 36•
LatitudeTM oil as a sustainable alternative to dietary fish oil in rainbow trout (Oncorhynchus mykiss): effects on filet fatty acid profiles, intestinal histology, and plasma biochemistryFront Sustain Food Syst, 62 (2022)
-
The first study on the performance of the Cargill/BASF EPA Canola oil in aquafeed studies on salmon.
- 37••
Genetically modified plants are an alternative to oily fish for providing n-3 polyunsaturated fatty acids in the human diet: a summary of the findings of a Biotechnology and Biological Sciences Research Council funded projectNutr Bull, 46 (2021), pp. 60-68, 10.1111/nbu.12478
-
An important demonstration of the utility of EPA + DHA Camelina oil for direct human nutrition. This confirms the ability of this novel oil to substitute for fish oils in direct human nutrition.
- 38
Canola engineered with a microalgal polyketide synthase-like system produces oil enriched in docosahexaenoic acidNat Biotechnol, 34 (2016), pp. 881-887, 10.1038/nbt.3585
- 39
Engineering of tomato for the sustainable production of ketocarotenoids and its evaluation in aquaculture feedProc Natl Acad Sci U S A, 114 (2017), Article 10876, 10.1073/pnas.1708349114–10881
- 40
Towards the development of a sustainable soya bean-based feedstock for aquaculturePlant Biotechnology Journal, 15 (2017), pp. 227-236, 10.1111/pbi.12608
- 41•
CRISPR-Cas9-Mediated gene editing of MYB28 genes impair glucoraphanin accumulation of Brassica oleracea in the fieldThe CRISPR journal, 4 (2021), pp. 416-426, 10.1089/crispr.2021.0007
-
Field-based demonstration of trait improvement by gene editing
- 42
Ablation of glucosinolate accumulation in the oil crop Camelina sativa by targeted mutagenesis of genes encoding the transporters GTR1 and GTR2 and regulators of biosynthesis MYB28 and MYB29Plant Biotechnol J (2022), 10.1111/pbi.13936
- 43
Nutritional enhancement in plants – green and greenerCurr Opin Biotechnol, 61 (2020), pp. 122-127, 10.1016/j.copbio.2019.12.010
- 44••
Systems approach to quantify the global omega-3 fatty acid cycleNat Food, 1 (2020), pp. 59-62, 10.1038/s43016-019-0006-0
-
An important framework for considering the volume of omega-3 fish oils available to the growing population and how best to close the gap between supply and demand. Current supplies can only provide <40% of demand.
- 45
Microbial and genetically engineered oils as replacements for fish oil in aquaculture feedsBiotechnol Lett, 39 (2017), pp. 1599-1609, 10.1007/s10529-017-2402-6
- 46
Microalgae oil as an effective alternative source of EPA and DHA for gilthead seabream (Sparus aurata) aquacultureAnimals, 11 (2021), p. 971, 10.3390/ani11040971Published 2021 Mar 31
- 47
Are fish what they eat? A fatty acid’s perspectiveProg Lipid Res, 80 (2020), Article 101064
- 48
A science-informed ethics for agricultural biotechnologyCrop Breeding, Genetics and Genomics, 1 (2019 Jul 19)



